上海有机所交叉中心揭示了程序性细胞死亡的广谱启动机制
程序性细胞死亡主要包括凋亡、程序性坏死、焦亡。凋亡由一系列caspases 蛋白介导,焦亡主要由caspases和gasdermin蛋白介导,而坏死通常发生在caspases被抑制的情况下。因此,不同的程序性细胞死亡具有相同/类似的分子特征。然而这些程序性死亡是否通过相同或类似的机制来启动仍然未知。
肿瘤坏死因子TNF是一种主要的促炎细胞因子,受体互作蛋白激酶RIPK1是TNF通路中的关键节点。RIPK1可以通过其激酶功能诱导凋亡和/或程序性坏死。正常生理情况下,RIPK1激酶活性被多个保护性的“刹车”,即细胞死亡检查点,所抑制,从而避免细胞死亡的发生。这些检查点由一系列复杂的翻译后修饰,主要包括泛素化和磷酸化,共同决定细胞是否发生死亡。任何一个检查点的缺失,都会使得RIPK1被激活,进而导致细胞死亡的发生。多种人类炎症性和自身免疫性疾病都与检查点缺失和RIPK1被激活有关。近年来,领域内已经鉴定出了多个RIPK1的保护性检查点。然而,当这些检查点失活时,RIPK1的激活以及细胞死亡的启动是否需要特定的信号仍不清楚。
2024年10月28日,中国科学院上海有机化学研究所生物与化学交叉研究中心许代超课题组在Molecular Cell杂志在线发表了题为“Palmitoylation licenses RIPK1 kinase activity and cytotoxicity in the TNF pathway”的研究论文,报道了RIPK1介导细胞死亡的启动机制,揭示了该机制以泛素化依赖的棕榈酰化的形式调控RIPK1的激活和程序性细胞死亡的分子机制。

S-棕榈酰化是一种可逆的脂质修饰,其特点是在棕榈酰基转移酶的催化下,棕榈酸通过硫酯键共价连接到蛋白的半胱氨酸残基。棕榈酰化可赋予蛋白疏水特性,促进蛋白的向膜迁移或影响蛋白互作。该研究发现,在TNF刺激短时间内可诱导RIPK1发生棕榈酰化修饰。修饰位点位于其激酶结构域内一个保守的半胱氨酸残基C257。进一步研究发现DHHC5是介导RIPK1棕榈酰化的主要棕榈酰基转移酶,且DHHC5的功能发挥依赖于RIPK1的K63泛素化对其进行的招募(图1)。
现有的理论认为,RIPK1的激活通过二聚体内的反式自磷酸化进行,这意味着其激酶结构域需要发生同源相互作用。然而,目前还不清楚RIPK1激酶结构域如何实现这种同源互作以促进自身磷酸化激活。该研究结果建立了以下模型详细阐明了当细胞死亡检查点被破坏时,RIPK1介导的细胞死亡如何被启动:①当细胞感应到TNF后,RIPK1被迅速招募到临近细胞膜区域的TNF受体复合物;在该复合物中,RIPK1在E3酶cIAP1/2的作用下发生K63连接的泛素化修饰。②这些K63泛素链会招募定位于细胞膜上的DHHC5,使之靠近RIPK1。③随后,DHHC5催化RIPK1激酶结构域上的棕榈酰化,增加激酶结构域的疏水特性,促进其同源相互作用;④在细胞死亡检查点受阻的情况下,上述过程可以增强RIPK1的反式自激活,最终导致下游的RIPK1介导的细胞死亡,包括凋亡和程序性坏死(图1)。
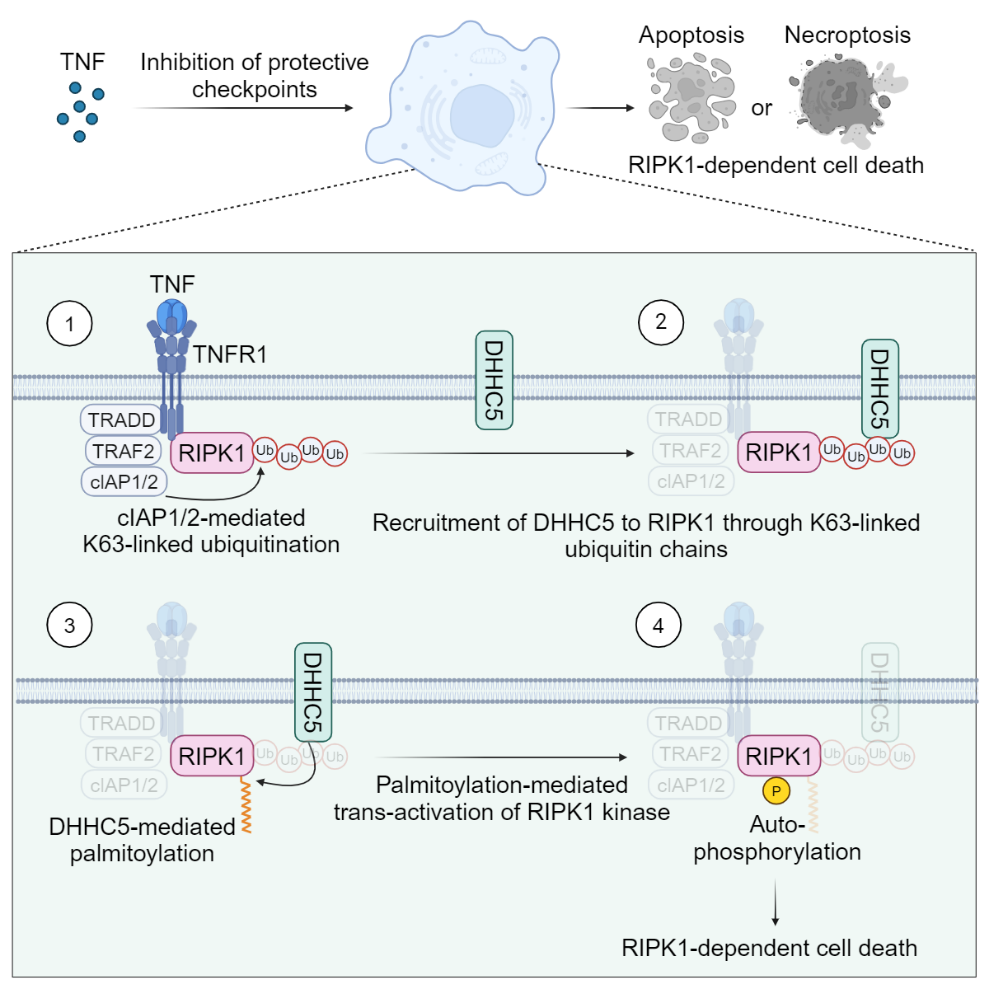
图1. 当TNF通路中的保护性检查点失活时,K63泛素化依赖的棕榈酰化促进RIPK1激活和启动细胞死亡的分子机制模式图。
该研究进一步表明这种修饰在体内至关重要,因为阻断RIPK1棕榈酰化可以保护小鼠免受TNF诱导的致死性休克。此外,在患有代谢功能障碍相关的脂肪性肝病(MASH)小鼠肝脏中,脂肪酸的累积促进了MASH肝脏中DHHC5的扩增和RIPK1的棕榈酰化,使得RIPK1过度激活导致肝细胞死亡和肝损伤。因此,该研究表明DHHC5介导的RIPK1棕榈酰化是治疗炎症性疾病的潜在靶点。
值得注意的是,许代超团队之前的研究发现gasdermin D的棕榈酰化是焦亡启动的必要条件(Nat Cell Biol 2024,26:757)。但棕榈酰化在不同细胞死亡类型中的作用方式有所不同。在焦亡中棕榈酰化促进gasdermin D的剪切激活和上膜成孔,而在凋亡和程序性坏死中,棕榈酰化促进RIPK1的反式自激活。总之,这些系列的研究表明棕榈酰化修饰是一种广谱的程序性细胞死亡启动机制。
中国科学院生物与化学交叉研究中心许代超研究员为本文通讯作者,华中科技大学同济医学院顾劲杨教授为共同通讯作者,中国科学院生物与化学交叉研究中心张娜博士为第一作者。中国科学院生物与化学交叉研究中心李婉津研究员、刘建平副研究员等人为该工作提供了宝贵的帮助。
原文链接:https://doi.org/10.1016/j.molcel.2024.10.002
附件下载: